Actin protein (>95% pure): rabbit skeletal muscle
Product Uses Include
- Identification and characterization of muscle actin binding proteins
- In vitro actin polymerization studies
- Antibody standard for Western blot analysis
Material
Actin protein has been purified from rabbit skeletal muscle. AKL99 actin is greater than 99% pure and AKL95 is greater than 95% pure. Muscle actin has an approximate molecular weight of 43 kDa. Rabbit muscle actin is supplied as a white lyophilized powder. The lyophilized protein when stored desiccated to < 10% humidity at 4°C is stable for 6 months. When re constituted in distilled water to 10 mg/ml, the protein is in the following buffer: 5 mM Tris-HCl pH 8.0, 0.2 mM CaCl2, 0.2 mM ATP, 5% sucrose, and 1% dextran.
Purity
Protein purity is determined by scanning densitometry of Coomassie Blue stained protein on a 12% polyacrylamide gel. AKL99 consists of >99% pure muscle actin while AKL95 is >95% pure (see Figure 1).
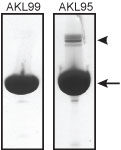
Figure 1: Figure 1. Purities of rabbit skeletal muscle actin protein. 100 µg of >99% pure (AKL99) and >95% pure (AKL95) rabbit skeletal muscle actin were run on SDS-PAGE gels and stained with coomassie blue. The arrow indicates actin protein, the arrowhead an α-actinin contaminant (115 kDa). The minor impurities in the purified actins are predominantly actin binding proteins such as α-actinin and gelsolin.
Biological Activity
The biological activity of muscle actinis determined by its ability to efficiently polymerize into filaments (F-actin) in vitro and separate from unpolymerized components in a spin down assay. Stringent quality control ensures that AKL99 produces > 90% F-actin and AKL95 produces > 80% F-actin in this assay.
For product Datasheets and MSDSs please click on the PDF links below. For additional information, click on the FAQs tab above or contact our Technical Support department at tservice@cytoskeleton.com
Question 1: What is the best way to store actin proteins to insure maximum stability and shelf-life?
Answer 1: Cytoskeleton provides all of our actin proteins as lyophilized powders so that they can be shipped at room temperature. Upon receipt, the lyophilized powders should be stored at 4°C in a sealed container with desiccant. It is important to monitor the freshness of the desiccant and insure that it continues to absorb moisture to protect the lyophilized actins. With proper storage, the lyophilized actins are guaranteed to be stable for 6 months from the date of purchase. Alternatively, actins can be immediately resuspended at the concentration recommended, aliquoted, snap-frozen in liquid nitrogen and stored at -70°C. The frozen aliquots will be stable for 6 months. When thawing frozen aliquots, it is important to thaw in a room temperature water bath.
Question 2: What is the best way to store F-actin after polymerizing?
Answer 2: G-actin is stable for two days at 4°C and requires a divalent cation, pH 6.5 - 8.0 and ATP for stability. F-actin is stable and can be stored at 4°C for 1-2 weeks. F-actin requires ATP (0.2 mM) and Mg2+ (2 mM) for stability and is unstable below pH 6.5 and above pH 8.5. F-actin is not stable to freezing. F-actin can be transferred to a variety of buffers (e.g. HEPES, phosphate, etc) without detrimental effects. We recommend the addition of antibacterial agents such as 100 μg/ml ampicillin and 10 μg/ml chloramphenicol when storing F-actin at 4°C.
If you have any questions concerning this product, please contact our Technical Service department at tservice@cytoskeleton.com.