Product Uses Include
- Fluorescent staining of fixed actin filaments in tissue sections and tissue culture cell preparations.Note: Unlike many actin antibodies, Acti-stain™ 670 phalloidin binds only to F-actin resulting in low background fluorescence. Furthermore, actin staining is not appreciably different between species.
- Preparation of stabilized fluorescent actin filaments in vitro.
Actin staining is very useful in determining the structure and function of the cytoskeleton in living and fixed cells. The actin cytoskeleton is a very dynamic and labile structure in the living cell, but it can be fixed with paraformaldehyde prior to probing or staining for actin structures.
Material
Phalloidin is a seven amino acid peptide toxin from the mushroom Amanita phalloides, which binds specifically and with high affinity (Kd 20 nM) to the polymerized form of actin (F-actin). Phalloidin lowers the critical concentration of actin polymerization to less than 1 µg/ml, thereby acting as a polymerization enhancer. Phalloidin has been labeled with a proprietary far-red fluorescent dye which allows it to be used to stain actin filaments in tissue cultured cells and tissue sections (1, see Figure 1) and cell-free preparations. Acti-stain™ 670 phalloidin-labeled actin filaments retain many functional characteristics of unlabeled actin including their ability to interact with myosin. Actin-stain™ 670 phalloidin is supplied as a light blue solid.
Note: Phalloidin is toxic and must be handled with care (LD50 human = 2mg/Kg).
Example Results and Specifications
Figure 1. Actin Stress Fibers stained with Acti-stain™ 670 in a Swiss 3T3 cell.
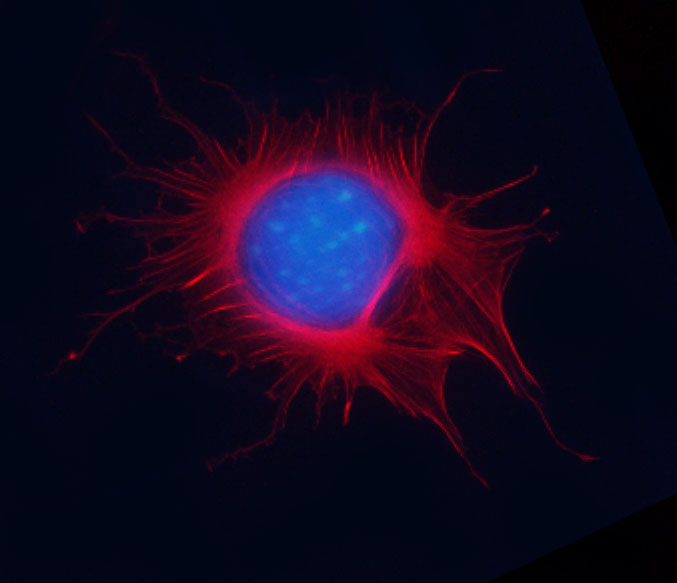
Mitotic Swiss 3T3 cell, F-actin stained with Acti-stain™ 670 (far-red, Cat. # PHDN1), nuclear DNA stained with Dapi (blue).
Figure 2. Emission and excitation scans for Acti-stain™ phalloidins
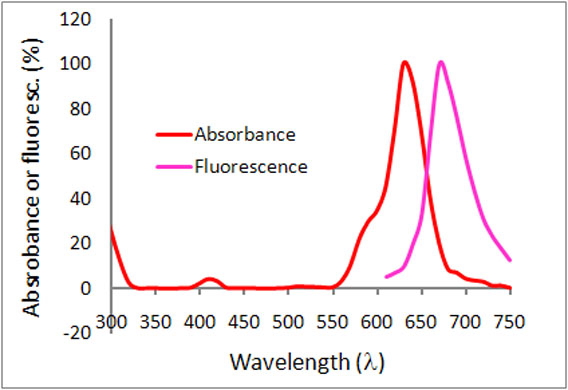
Absorbance and fluorescence scan of Acti-stain™ 670. Labeled phalloidin was diluted into methanol and its absorbance and excitation spectra were scanned between 300-750 and 600-750 nm, respectively. Absorbance peaks at 625 nm and fluorescence at 675 nm.
Acti-stain phalloidins are the most well characterized phalloidins available. Tabe 1 describes their brightness, photostability, background and affinity constants to F-actin. Compare these performance characteristics to other fluorescent phalloidins and you will see the advantages of using Acti-stain™ for your actin staining requirements.
Table 1. Biochemical characteristics of fluorescent phalloidins
| Conjugate | Cat.# | Wavelengths (Ex/Em) | Brightness (AFU*) | Stability to photobleaching(half life in seconds**) | Background (% of total AFU at 100nM**) | Affinity (Kd in nanomolar) |
Fluorescein-phalloidin | na | 485/535 FITC filter set | 432 | 6 | 22 | 72 +/-12 |
Acti-stain™ 488 | PHDG1 | 485/535 FITC filter set | 832 | 57 | 5 | 55 +/-8 |
Acti-stain™ 535 | PHDR1 | 535/585 TRITC filter set | 430 | 27 | 12 | 36 +/-7 |
Acti-stain™ 555 | PHDH1 | 535/585 TRITC filter set | 551 | 46 | 16 | 63 +/-8 |
Acti-stain™ 670 | PHDN1 | 640/680 Cy5 filter set | 332 | 18 | 18 | 50 +/-12 |
* = AFU's measured by quantitative cell imaging. ** = Measured in stained Swiss 3T3 cells in the absence of antifade.
For more information about protocols to fix and stain cells click here.
References
1. Wulf, E. et al. (1979). Proc Natl Acad Sci USA. 76(9):4498-4502.
2. Kron, S.J. et al. (1991). Meth. Enzmol. 196: 399-416.
For product Datasheets and MSDSs please click on the PDF links below. For additional information, click on the FAQs tab above or contact our Technical Support department at tservice@cytoskeleton.com
Question 1: Can I use fluorescently-labeled phalloidin to stain F-actin in living cells?
Answer 1: Unfortunately, no, phalloidin cannot be used to stain F-actin in living cells. Phalloidins are used to stain F-actin in fixed cells. Fluorescent phalloidins only bind to the native quaternary structure of F-actin which provides a low background. The correct fixation condition for phalloidin binding is 3.7% (v/v) paraformaldehyde in PBS for 10 min because it retains the quaternary protein structure which is necessary for high affinity binding of phalloidin. Methanol fixation destroys the native conformation and hence is not suitable for F-actin staining with phalloidin. To monitor actin dynamics in living cells, micro-injection of rhodamine-labeled actin (Cat. # APHR or AR05) is recommended. Please see those datasheets for more information.
Question 2: Which of the fluorescently-labeled phalloidin is the most stable/brightest?
Answer 2: The brightest and most stable of the Acti-stains is Acti-stain 488 (green fluorescence; Cat. # PHDG1). Please see the table below for additional information on all of our Acti-stains.
Conjugate | Cat. # | Wavelengths (Ex/EM) | Brightness (AFU) | Stability to photobleaching* (1/2 life in sec) | Background (% of total AFU at 100 nM) |
Acti-stain™ 488 | 485/535 | 832 | 57 | 5 | |
Acti-stain™ 535 | 535/585 | 430 | 27 | 12 | |
Acti-stain™ 555 | 535/585 | 551 | 46 | 16 | |
Acti-stain™ 670 | 640/680 | 332 | 18 | 18 |
* = measured in the absence of antifade.
If you have any questions concerning this product, please contact our Technical Service department at tservice@cytoskeleton.com.