October 2020 Newsletter : Coronavirus and the Cytoskeleton
The Coronavirus superfamily (Coronaviridae) includes several human pathogens with large positive stranded RNA-encoded genomes, e.g. common cold, viral encephalitis and Covid-19, which are classified into alpha-, beta-, gamma- and delta-coronavirus families by phylogenetic clustering, with further division into Lineages A, B, and C. The virus that causes COVID-19 is classed as a lineage B beta-coronavirus with high similarity to SARS-CoV, thus it's renaming to SARS-CoV-2 (1). Although the first members of the beta-coronavirus family were recorded in the 1960’s, the family’s rate of new virulent human pathogens has increased rapidly over the past 20 years, now numbering six in total with the latest ones having the familiar pseudonyms SARS (Severe Acute Respiratory Syndrome, 2002), HKU1 (HongKong, 2005), MERS (Middle East Respiratory Syndrome, 2012), and now COVID-19. Their emergence is linked to increased density of human and animal populations which has enhanced zoonotic transmission rates (2).
Here, we describe the five stages of virus “life” cycle with respect to its interaction with the cytoskeleton. The first stage of infection for Coronaviridae is Spike (S) protein-mediated attachment to the cell surface via its binding to a prevalent receptor called angiotensin-converting enzyme 2 (ACE2). The receptor is widespread in air-track and lung tissue, and is more prevalent in older males which accounts for their higher death rates.
After binding, virus particles actively rearrange the cytoskeleton by regulating the FAK/Cofilin/Rac/Cdc42 pathway (4). Lv et al. showed temporal fluctuations in filopodia, lamellipodia, and stress fiber proportions after administering coronavirus PHEV to mouse brain neuronal (N2a) cells in vitro. Interestingly, within 5 min, stress fibers were largely depleted, whereas filopodia dominated the detectable F-actin content, producing 80% of the total fluorescent phalloidin signal (4). Between 20-40 min, lamelopodia emerged as an equal co-player, and finally at 60 min, proportions returned to near normal levels, i.e., 60% of the signal was stress fibers. Although this is a macroscopic observation, it indicates on a nanometer scale that virions may create a local environment that regulates their own individual cellular entry. In another report, Owczarek et al. administered coronavirus OC43 to human colorectal adenocarcinoma (HCT-8) cells to study the method of invagination, vesical excision, and function of F-actin (5). Clathrin-dependent endocytosis, caveolae- dependent invagination, and pinocytosis were all implicated in redundant virion entry mechanisms. And dynamin, a large GTPase, was necessary for vesical entry and excision (5). Similar studies of viral entry were described by Milewska et al. (6) with active endocytosis or mass action entry of virions through other mechanisms, e.g., caveolae- dependent invagination or pinocytosis, with the infection rate being dependent on the viral loading of the cell surface. Cellular entry also packages the virion for the second stage which is transportion to the perinuclear region (5).
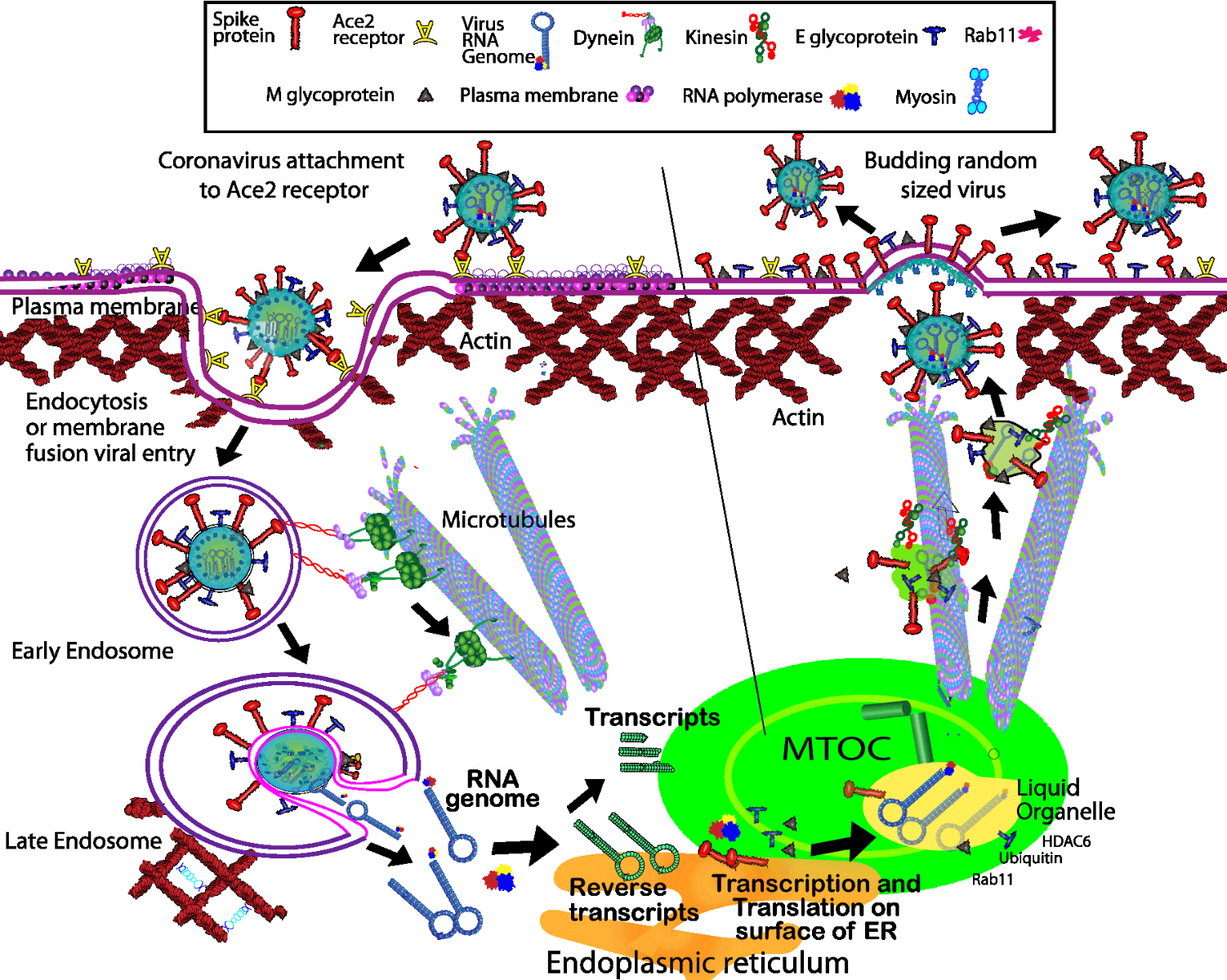
Figure Top: Schematic diagram of the current knowledge of coronavirus entry, replication, assembly, and egress from the host cell (adapted from Simpson & Yamauchi, 2020).
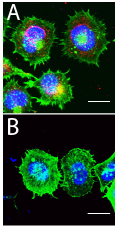
Figure Bottom: HCT-8 cells treated or untreated with PHEV coronavirus for 5 min, cells were fixed and stained with Acti-stain 488 (green, Cytoskeleton), anti-PHEV coat protein (red) and DNA (blue, Hoescht). Top - Infected with coronavirus, bottom - Un-infected. Note high level of staining on the periphery of the cell in the infected samples indicating lamellipodia and filopodia (narrow spikes). Compared to the un-infected cell which indicate a high level of cytosplasmic stianing indicating stress fibers. (Kindly provided by Prof. Wenqi He, from Lv et al. 2019).
Several studies have implicated both actin and tubulin based systems are complementary cytoskeleton components of intracellular transport. Owczarek et al. probed the function of F-actin in intracellular localization; interestingly, jasplakinolide, a cell permeable F-actin stabilizing compound, inhibited viral entry of plasma membrane bound virions, whereas cytochalsin D, an F-actin depolymerizing compound, did not inhibit viral entry but did disrupt the normal localization of virions from peri-nuclear to cytoplasmic areas (5). In contrast, Rüdiger et al. used a spin-down format to show preferential tubulin isoform binding of virions. Coronavirus Spike protein C-terminal peptide (S-protein) bound to several beta-tubulin isoforms in a coronavirus strain-specific manner (9). A scrambled peptide showed the binding was not due to random ionic charge interaction. Thus, intracellular transport of virions utilizes multiple cytoskeletal structural proteins to navigate through and localize to specific areas within the cell (see Figure 1).
The third stage is, transcription and translation. After transport to the endoplasmic reticulum(ER)/Golgi/microtubule organizing center(MTOC)/perinuclear (PN) region (see Figure 1), coronavirus RNA exudes from the endosome vesicle and virion capsule and is translated to form two contiguous multigene polypeptides, which contains proteases that clip the peptide into functional proteins. The positive strand RNA is also reverse transcibed by an RNA polymerase dependent reverse transcriptase to form an antisense replication template. This replicon is subsequently transcribed back into positive sense RNA which is the starting point for viroid assembly and maturation. All these processes occur in the Golgi/ER/MTOC/PN region which is why drugs that disrupt ER, Golgi or MTOC also inhibit viral replication and reduce viral load (10).
The fourth stage is assembly and maturation. Initially, the nucleocapsid (N) protein binds to an RNA copy and binds to vesicle membranes (10,11), and further maturation occurs with N and E proteins which are required for assembly of the basic virus-like particle (VLP). If Spike (S) protein is co-expressed, then this is incorporated into the virus particle (10). In concert with several cytoskeleton and membrane regulator proteins, e.g., HDAC6, ubiquitin, and Rab GTPases, which aid assembly by concentrating packaging components (7). It is unknown which one of these predominates for COVID-19 / SARS-CoV-2.
The fifth stage is egress. The genetic fusion of the coronavirus nucleocapsid or spike proteins with GFP permits tracking of non-infective virus particles by fluorescent microscopy. Using this technique, Siu et al. monitored SARS-CoV egress and, found vesicles that fused into multi-particle conglomerates (10). The transport was sensitive to Brefeldin A, which indicates that the secretory pathway was being used. Other studies have found nocodazole was an effective inhibitor of virion transport to the plasma membrane (7,12), indicating that microtubules are an essential component of egress. Rab11 is implicated in binding to KHC for microtubule transport and then binds to myosin to help transverse the peripheal actin matrix and egress from the cell.
In summary, coronavirus has a highly specific mechanism of receptor binding with multiple mechanisms to enter the cell. Subsequently, both actin, tubulin, and their chemo-mechanical motors dynein, kinesin and myosin cytoskeleton components are required for intracellular transport to the correct location for translation and replication. After reverse transcription and transcription, positive strand RNA is packaged on a scaffold of Golgi/ER/microtubule complexes. The vesicle-encased virions track along microtubules before fusing with the plasma membrane and escaping the cell. Many of the methods in coronavirus research use reagents such as fluorescent phalloidins, microtubule and actin spin-down kits, dynein, kinesin proteins, and Rac and Cdc42 activation assays that are present in the Cytoskeleton catalog and noted below in the tables. There are many un-answered questions in coronavirus research; for example, how do the virions regulate the cytoskeleton through the plasma membrane interactions to coordinate their entry? What is the significance of using actin and microtubule cytoskeletons for different functions? How does the virus evade the immune system for so long compared to influenza and the common cold? Answering these questions will undoubtedly lead to greater knowledge and possible pharmacological breakthroughs in the future.
References
1. Gorbalenya, A.E., Baker, S.C., Baric, R.S. et al. The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol (2020). https://doi.org/10.1038/s41564-020-0695-z.
2. Vijgen, L. et al. 2005. Complete genomic sequence of human coronavirus OC43: molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J Virol i79,i1595–1604, https://doi.org/10.1128/JVI.79.3.1595-1604.
3. Ren et al. 2015. Genetic drift of human coronavirusOC43 spike gene during adaptive evolution. Scientific Reports, 5, 11451.
4. Lv et al. 2019. Porcine hemagglutinating encephalomyelitis virus activation of the integrin a5b1-FAK-cofilin pathway causes cytoskeletal rearrangement to promote its invasion of N2a cells. J. Virol. 93 (5), 1-19. doi: org/10.1128/JVI.01736-18.
5. Owczarek et al. 2018. Early events during human coronavirus entry to the cell. Scientific Reports, 8, 7124, 1-11. doi: 10-1038/s41598-018-25640-0.
6. Milewska, A.et al. 2014. Human coronavirus NL63 utilizes heparan sulfate proteoglycans for attachment to target cells. J Virol 88, 13221–13230, https://doi.org/10.1128/JVI.02078-14.
7. Simpson C. and Yamaguchi Y., 2020. Microtubules in Influenza Virus Entry and Egress. Viruses 12(1), 117; https://doi.org/10.3390/v12010117.
8. Astuti, I. and Ysrafil, 2020. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2): An overview of viral structure and host repsonse.
9. Rüdiger et al 2016. Tubulins interact with porcine and human S proteins of the genus Alphacoronavirus and support successful assembly and release of infectious viral particles. Virology 497 (2016) 185–197. http://dx.doi.org/10.1016/j.virol.2016.07.022.
10. Siu et al., 2008. The M, E, and N Structural Proteins of the Severe Acute Respiratory Syndrome Coronavirus Are Required for Efficient Assembly, Trafficking, and Release of Virus-Like Particles. J. Virol. p. 11318–11330. doi:10.1128/JVI.01052-08
11. Hsin et al. 2018. Nucleocapsid protein-dependent assembly of the RNA packaging signal of Middle Eastern respiratory syndrome coronavirus. J. Biomed. Sci. 25(47), 1-12. doi.org/10.1186/s12929-018-0449-x.
12. Ward, 2011. The taking of the cytoskeleton one two three: How viruses utilize the cytoskeleton during egress. Virology, 411 (2), 244-250. doi:10.1016/j.virol.2010.12.024.
Related Products
G-LISA Kits
Rac1,2,3 G-LISA Activation Assay (Colorimetric format) (Cat. # BK125)
Cdc42 G-LISA Activation Assay (Colorimetric format) (Cat. # BK127)
Rac1 G-LISA Activation Assay Kit (Colorimetric Based) (Cat. # BK128)
Acti-Stain Phalloidins
Acti-stain 488 Phalloidin (Cat. # PHDG1)
Acti-stain 555 Phalloidin (Cat. # PHDH1)
Acti-stain 670 Phalloidin (Cat. # PHDN1)
Rhodamine Phalloidin (Cat. # PHDR1)