April 2020 Newsletter: F-Actin Probes in Living Cells
F-Actin Probes in Living Cells
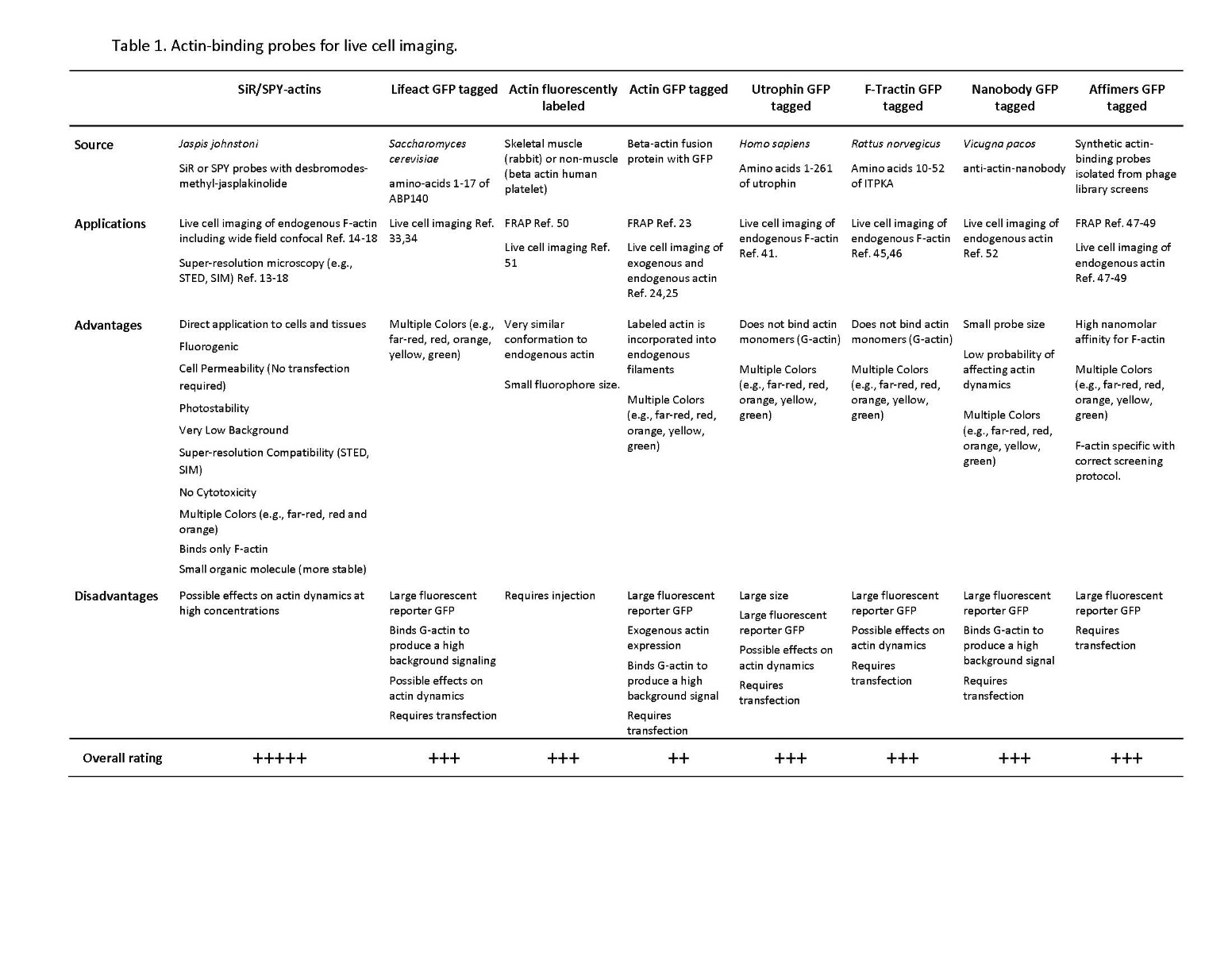
Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)] is required for multiple physiological functions, including intracellular transport, cell growth, morphology, motility, trafficking, polarity, cell-to-cell contacts, and cytokinesis1,2. Correspondingly, dysfunctional actin cytoskeletal dynamics are a pathophysiological feature of many human diseases, including those with oncogenic, neurodegenerative, or cardiovascular origins3-9. For these reasons, there is continuing interest in F-actin live cell imaging probes to study actin cytoskeletal dynamics in cell culture models of health and disease (Table 1).
SiR and SPY Actin Probes
The ideal actin visualization tool is a small molecule able to bind F-actin in a sensitive and selective manner, while not disrupting actin re-modeling. In addition, introduction directly into the cell culture medium or tissues without need for transfection or electroporation is advantageous10-13. The new SiR/SPY actin probes fulfill the needs of an “ideal” actin-binding molecule while surmounting most, if not all, of the concerns and shortcomings associated with existing actin probes (Table 1; Figs. 1,2). Initially characterized by Lukinavicius et al.12,13 and introduced commercially in 2014, the SiR and SiR700-actin live cell imaging probes label endogenous F-actin and avoid the need for transfections and over-expression of labeled actin proteins or actin-binding proteins 12,13. SiR/SiR700-actin probes are structurally related to the naturally-occurring F-actin binding molecule jasplakinolide12,13. These F-actin probes utilize the proprietary fluorophore silicon rhodamine (SiR), a bright, photostable far-red dye with little, if any, background signal (Figs. 1,2). Because SiR probes exist in a closed, non-fluorescent state (spirolactone), the probes are self-quenching when unbound to F-actin12,13 (Fig. 3). SiR probes are visualized with standard Cy5 settings (optimal excitation, 650 nm; emission, 670 nm) which confer compatibility with a wide range of genetically-encoded reporter fluorophores (e.g., GFP, m-Cherry)12,13. SPY555-actin is the newest addition to Spirochrome’s family of F-actin live cell imaging probes. SPY555-actin is an improved version of the SiR-actin probes as a lower concentration can be used which offers robust labeling and reduced cytotoxicity and perturbation of actin cytoskeletal dynamics. SPY555-actin is imaged with a standard TMR or Cy3 channel (optimal excitation, 555 nm; emission, 580 nm) using the same staining protocol as for SiR/SiR700-actins. The key features of SiR and SPY actin probes are their cell permeability, fluorogenic character, minimal cytotoxicity, photostability, and compatibility with both standard fluorescence microscopy (e.g., wide-field, confocal) and super-resolution microscopy (e.g., STED, SIM)12-18 (Figs. 1,2). The combination of STED and SiR/SPY-actin probes allows for unparalleled fluorescent visualization of subcellular F-actin structures and their physical characterization in living cells14-18 (Figs. 1,2). SiR-actin probes have been used to examine F-actin in tissue19 and a wide variety of cell types, including (but not limited to) human-induced pluripotent stem cell lines, cardiac cells, endothelial cells, epithelial cells, muscle cells, multiple cancer cell lines, and primary neurons 14,16-18,20-22 (Figs. 1,2).


Figure 1. MCF10A cells expressing H2B-GFP (blue) in Matrigel (3D culture) stained with SiR-actin (red). Image taken on an inverted LSM microscope. Courtesy of Christian Conrad and Katharina Jechow, Heidelberg.
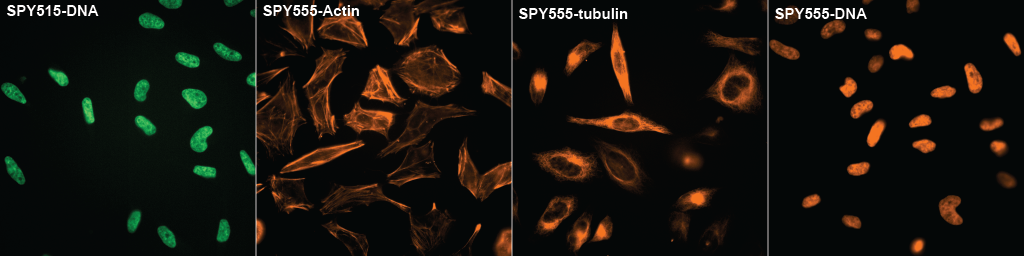
Figure 2. SPY505 and SPY555 staining DNA,Actin,Tubulin, and DNA . Photo comes from front page of Spirochrome's website.
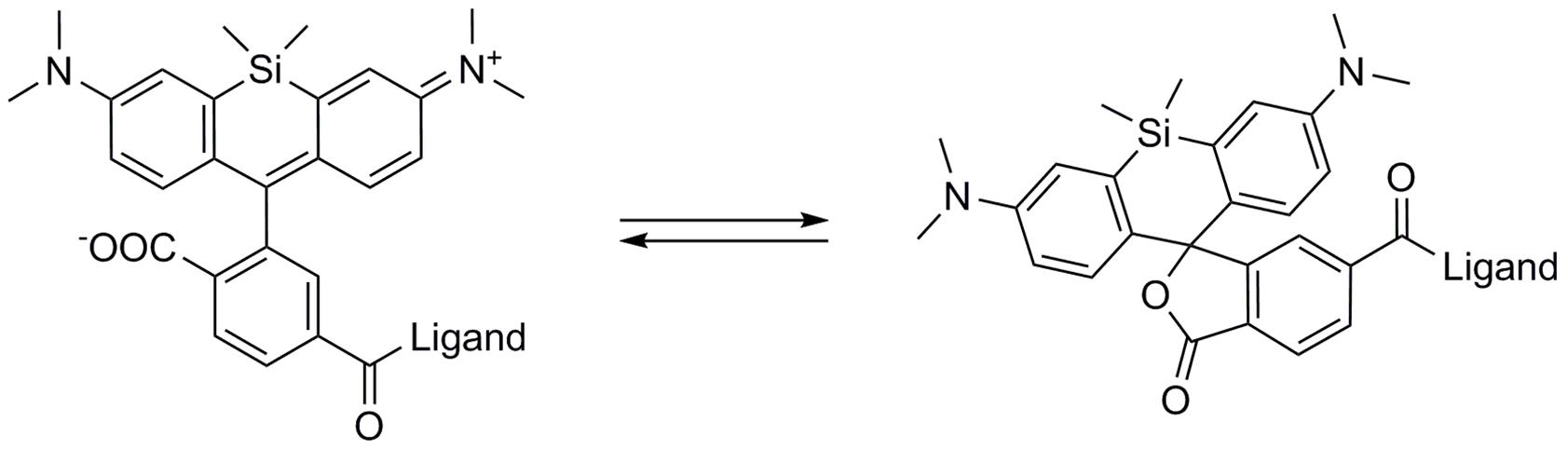
Figure 3. SiR derivatives exist in equilibrium between the fluorescent zwitterionic (open) form (left structure) and the non-fluorescent spiro (closed) form (right structure).
Fluorescent actin and fluorescent actin-binding domains
The first studies of live cell actin dynamics were performed with fluorescent derivatives of actin protein whch were microinjected into cells50,51. This was a highly effective procedure but the apparatus took a while to setup. Thus overtime, transfections of GFP-actin conjugates became more popular. Fluorescently labeled actin protein or GFP/eGFP-actins worked well with fluorescence recovery after photobleaching (FRAP) microscopy11,23-25.which indicates the dynamic nature of actin cytoskeleton rearrangements. However, GFP/eGFP-actin has several drawbacks10. First, the size of GFP (~28 kDa) can impair polymerization26 and GFP-actin can differentially label F-actin structures10,24. Second, some actin-binding proteins (e.g., formin family nucleators) might sterically hinder incorporation of GFP-actin into actin seeds or growing polymers27,28. Third, there is a relatively high background signal from non-filamentous fluorescent actin29. Finally, expression of eGFP-actin can affect cell behavior30,31.
Another method for actin live cell imaging utilizes yeast- or human-derived actin binding domains fused to GFP, eGFP, or m-Cherry fluorophores10,11,32. The most common genetically-encoded F-actin probes are Lifeact, utrophin (UtrCH), and F-tractin10. Lifeact is a 17 amino acid peptide from yeast Abp14033,34 used for live cell imaging in mammalian and non-mammalian cells 24,34-36. Lifeact has several disadvantages, including the possibility of affecting actin dynamics (so-called Lifeact-induced artifacts) and inhibiting the binding of actin-associated proteins such as cofilin32,37-40. Although Lifeact-GFP binds strongly to F-actin (Kd, 2.2 ± 0.3 μM), its binding affinity for G-actin is 10-fold higher33, resulting in high background fluorescence. Lifeact does not bind all actin-containing structures10,38. Lifeact is introduced into the cell through transfection rather than simply adding it into the medium as is done for the SiR/SiR700/SPY probes. UtrCH is based on the tandem calponin homology domains (CH1 and CH2) of utrophin41 and consists of the first 261 amino acid residues of human utrophin, an actin binding protein42. The CH domains bind to actin with a Kd of ~18 μM43. Utrophin-based probes have been used successfully across a wide range of cell types and species10,32. Similar to Lifeact, at high concentrations, utrophin-based probes can exert deleterious effects on actin cytoskeletal dynamics32,37,44. F-tractin is a 43 amino acid peptide derived from the rat actin-binding inositol 1,4,5-triphosphate 3-kinase A which binds F-actin with a Kd of ~10 μM45,46. Due to its larger size (in comparison to other probes), F-tractin might sterically hinder binding of actin-binding proteins that regulate and/or facilitate polymerization10 and can modify actin-based cellular structures32.
Actin-directed nanobodies and affimer proteins for F-actin
Two new technologies for monitoring actin dynamics in living cells are 1. single-domain antibodies, so-called nanobodies52, and 2. actin “affimers” - synthetic, actin-binding proteins isolated from phage library screens47-49. If not developed correctly, nanobodies can exhibit a high background signal due to G-actin binding10. Recently, three eGFP-fusion actin affimers were described with low micromolar binding affinities for F-actin47, but FRAP microscopy suggests that the eGFP-affimers may preferentially bind to a subset of actin filaments and alter actin organization in the cell47.
Summary
Despite multiple options for visualizing F-actin-based structures in living cells, there is no perfect live cell probe (Table 1). Ideally, the best F-actin probe will be sensitive, selective, fluorogenic, produce a very low background signal, non-toxic, and easily introduced into a wide range of cells across multiple species. It is of paramount importance to confirm that changes in actin cytoskeleton dynamics/structural organization are physiologically relevant and not artifacts of the probe itself. To assist researchers in F-actin live cell imaging studies, Cytoskeleton offers the SiR and SPY actin live cell imaging probes.
References
- Pollard T.D. 2016. What we know and do not know about actin. In: Jockusch B. (eds.) The Actin Cytoskeleton. Handbook of Experimental Pharmacology. vol 235. Springer, Cham.
- Dominguez R. and Holmes K.C. 2011. Actin structure and function. Annu. Rev. Biophys. 40, 169-186.
- Verstraelen P. et al. 2018. Image-based profiling of synaptic connectivity in primary neuronal cell culture. Front. Neurosci. 12, 389.
- Hoover B.R. et al. 2010. Tau mislocalization to dendritic spines mediates synaptic dysfunction independently of neurodegeneration. Neuron. 68, 1067-1081
- Spires-Jones T.L. and Hyman B.T. 2014. The intersection of amyloid beta and tau at synapses in Alzheimer’s disease. Neuron. 82, 756-771.
- Dugina V.B. et al. 2019. Biological role of actin isoforms in mammalian cells. Biochemistry (Mosc). 84, 583-592.
- Hartmann S. et al. 2015. The function of Rho-associated kinases ROCK1 and ROCK2 in the pathogenesis of cardiovascular disease. Front. Pharmacol. 6, 276.
- Bonello T.T. et al. 2009. New approaches to targeting the actin cytoskeleton for chemotherapy. Future Med. Chem. 1, 1311-1331.
- Bryce N.S. et al. 2019. Chemical biology approaches targeting the actin cytoskeleton through phenotypic screening. Curr. Opin. Chem. Biol. 51, 40-47.
- Melak M. et al. 2017. Actin visualization at a glance. J. Cell Sci. 130, 525-530.
- Belin B.J. et al. 2014. Comparative analysis of tools for live cell imaging of actin network architecture. Bioarchitecture. 4, 189-202.
- Lukinavicius G. et al. 2013. A near-infrared fluorophore for live-cell super-resolution microscopy of cellular proteins. Nat. Chem. 132-139.
- Lukinavicius G. et al. 2014. Fluorogenic probes for live-cell imaging of the cytoskeleton. Nat. Methods. 11, 731-733.
- D’Este E. et al. 2015. STED nanoscopy reveals the ubiquity of subcortical cytoskeleton periodicity in living neurons. Cell Rep. 10, 1246-1251.
- Hell S.W. and Wichmann J. 1994. Breaking the diffraction resolution limit by stimulated emission: stimulated-emission-depletion fluorescence microscopy. Opt. Lett. 19, 780-782.
- Bar J. et al. 2016. Periodic F-actin structures shape the neck of dendritic spines. Sci. Rep. 6, 37136.
- Liete S.C. et al. 2016. The actin-binding protein α-adducin is required for maintaining axon diameter. Cell Rep. 15, 490-498.
- Wang L. et al. 2019. Small-molecule fluorescent probes for live-cell super-resolution microscopy. J. Am. Chem. Soc. 141, 2770-2781.
- Klementieva N.V. et al. 2016. Fluorescence imaging of actin fine structure in tumor tissues using SiR-actin staining. Anticancer Res. 36, 5287-5294.
- Kim T.H. et al. 2016. Cancer cells become less deformable and more invasive with activation of β-adrenergic signaling. J. Cell Sci. 129, 4563-4575.
- Hashimoto N. et al. 2017. Reversible differentiation of immortalized human bladder smooth muscle cells accompanied by actin bundle reorganization. PLoS ONE. 12, e0186584.
- Narva E. et al. 2017. A strong contractile actin fence and large adhesions direct human pluripotent colony morphology and adhesion. Stem Cell Reports. 9, 67-76.
- Choidas A. et al. 1998. The suitability and application of a GFP-actin fusion protein for long-term imaging of the organization and dynamics of the cytoskeleton in mammalian cells. Eur. J. Cell Biol. 77, 81-90.
- Lemieux M.G. et al. 2014. Visualization of the actin cytoskeleton: different F-actin-binding probes tell different stories. Cytoskeleton. 71, 157-169.
- Small J. et al. 1999. Visualising the actin cytoskeleton. Microsc. Res. Tech. 47, 3–17.
- Sliogeryte K. et al. 2016. Differential effects of LifeAct-GFP and actin-GFP on cell mechanics assessed using micropipette aspiration. J. Biomech. 49, 310-317.
- Wu J.-Q. and Pollard T.D. 2005. Counting cytokinesis proteins globally and locally in fission yeast. Science. 310, 310-314.
- Chen Q. et al. 2012. Formins filter modified actin subunits during processive elongation. J. Struct. Biol. 177, 32-39.
- Lee C.W. et al. 2013. Dynamic localization of G-actin during membrane protrusion in neuronal motility. Curr. Biol. 23, 1046–1056.
- Aizawa H. et al. 1997. A green fluorescent protein-actin fusion protein dominantly inhibits cytokinesis, cell spreading, and locomotion in Dictyostelium. Cell Struct. Funct. 22, 335–345.
- Nagasaki A. et al. 2017. The position of the GFP tag on actin affects the filament formation in mammalian cells. Cell Struct. Funct. 42, 131–140.
- Patel S. et al. 2017. Functional characterisation of filamentous actin probe expression in neuronal cells. PLoS ONE. 12, e0187979.
- Riedl J. et al. 2008. Lifeact: a versatile marker to visualize F-actin. Nat. Methods. 5, 605-607.
- Riedl J. et al. 2010. Lifeact mice for studying F-actin dynamics. Nat. Methods. 7, 168-169.
- Schachtner H. et al. 2012. Tissue inducible Lifeact expression allows visualization of actin dynamics in vivo and ex vivo. Eur. J. Cell Biol. 91, 923-929.
- Mizoguchi T. et al. 2016. Zebrafish lines expressing UAS-driven red probes for monitoring cytoskeletal dynamics. Genesis. 54, 483-489.
- Du J. et al. 2015. Lifeact and Utr230 induce distinct actin assemblies in cell nuclei. Cytoskeleton. 72, 570-575.
- Munsie L.N. et al. 2009. Lifeact cannot visualize some forms of stress-induced twisted F-actin. Nat. Methods. 6, 317.
- Courtemanche N. et al. 2016. Avoiding artefacts when counting polymerized actin in live cells with LifeAct fused to fluorescent proteins. Nat. Cell Biol. 18, 676-683.
- Flores L.R. et al. 2019. Lifeact-TagGFP2 alters F-actin organization, cellular morphology and biophysical behaviour. Sci. Rep. 9, 3241.
- Burkel B.M. et al. 2007. Versatile fluorescent probes for actin filaments based on the actin-binding domain of utrophin. Cell Motil. Cytoskelet. 64, 822-832.
- Winder S.J. et al. 1995. Calmodulin regulation of utrophin actin binding. Biochem. Soc. Trans. 23, 397S.
- Rybakova I.N. and Ervasti J.M. 2005. Identification of spectrin-like repeats required for high affinity utrophin-actin interaction. J. Biol. Chem. 280, 23018–23023.
- Spracklen A.J. et al. 2014. The pros and cons of common actin labeling tools for visualizing actin dynamics during Drosophila oogenesis. Dev. Biol. 393, 209-226.
- Schell M.J. et al. 2001. Inositol 1,4,5-triphosphate 3-kinase A associates with F-actin and dendritic spines via its N-terminus. J. Biol. Chem. 276, 37537-37546.
- Brehm M.A. et al. 2004. Identification of the actin-binding domain of Ins(1, 4, 5)P3 3-kinase isoform B (IP3K-B). Biochem. J. 382, 353–362.
- Lopata A. et al. 2018. Affimer proteins for F-actin: novel affinity reagents that label F-actin in live and fixed cells. Sci. Rep. 8, 6572.
- Tiede C. et al. 2014. Adhiron: a stable and versatile peptide display scaffold for molecular recognition applications. Protein. Eng. Des. Sel. 27,145–155.
- Tiede C. et al. 2017. Affimer proteins are versatile and renewable affinity reagents. Elife 6, 24903.
- Shimada Y. et al. 1997. Dynamics of actin in cardiac myofibrils and fibroblast strees fibers. Cell Structure and function, 22, 59-64.
- Machesky L.M. and Hall A. 1997. Role of actin polymerization and adhesion to extracellular matrix in Rac- and Rho-induced cytoskeletal reorganization. J. Cell Biol. 138, 913-926.
- Rocchetti A. et al. 2014. Fluorescent labelling of the actin cytoskeleton in plants using a cameloid antibody. Plant Methods 10, 12 10.1186/1746-4811-10-12.