Exoenzyme C3 transferase protein: His tagged:Clostridium botulinum recombinant
Product Uses Include
- Inhibition of RhoA, B and C in vitro
- Inhibition of RhoA, B and C in vivo (See Table 1 for examples of use)
Material
Exoenzyme C3 transferase is an ADP ribosyl transferase that selectively ribosylates RhoA, RhoB and RhoC proteins on asparagine residue 41, rendering them inactive. It has extremely low affinity for other members of the Rho family such as Cdc42 and Rac1 and does therefore not affect these GTPases. Hence, C3 transferase is a very potent and useful reagent to specifically block RhoA/B/C signaling.
C3 transferase has been produc ed by expression in E. coli as a His-tagged protein. The recombinant protein is 24 kDa in size and is supplied as a lyophilized powder. Reconstitution of the protein in water to 1 mg/ml leaves the protein in the following buffer: 20 mM Tris pH 7.5, 50 mM NaCl, 0.5% sucrose and 0.1% dextran. Protein concentration is determined by the Precision Red Advanced Protein Assay Reagent Cat. # ADV02.
C3 transferase protein is also available in a cell permeable format (Cat. CT04) for fast, efficient and simple inhibition of RhoA/B/C in living cells.
Purity
Purity is determined by scanning densitometry of protein run on SDS-PAGE gels. CT03 consists of more than 90% pure exoenzyme C3 Transferase.
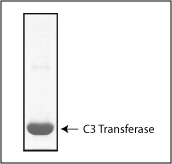
Figure 1: Exoenzyme C3 transferase purity determination. A 20 µg sample of CT03 was separated by SDS-PAGE and protein was stained with coomassie blue. Protein quantitation was performed using Precision Red Assay reagent (Cat. # ADV02). Purity was determined by scanning densitometry. The protein was determined to be >90% pure.
Biological Activity
Biological activity of C3 transferase is verified by the ability of the protein to ribosylate RhoA protein in platelet lysates in vitro (Fig. 2).
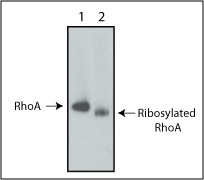
Figure 1: ADP-ribosylation of RhoA protein in human platelet extract. Platelet extract (100 µg) was reacted with C3 protein (1 µg) for 30 min at 37°C. Extracts were run on non-denaturing gel electrophoresis and RhoA protein was detected by Western blot. Lane 1 shows untreated extracts. Lane 2 shows C3 transferase treated extracts. The RhoA in the C3 transferase treated extract shows increased migration in the gel due to its ADP ribosylation.
| Table 1. Examples of how Cytoskeleton, Inc's C3 transferase has been used on cells to inactivate RhoA. | |||
Cell type | Method of introduction into cells | Concentration of C3 used (mg/ml) | Citation |
BS-C-1 | microinjection | 0.1 | 2 |
NRK | added to culture media | 0.02 | 7 |
Xenopus oocytes | microinjection | 0.08 | 1 |
Primary Aplysia bag cells | microinjection | 0.3 | 8 |
For product Datasheets and MSDSs please click on the PDF links below. For additional information, click on the FAQs tab above or contact our Technical Support department at tservice@cytoskeleton.com
Question 1: What is the difference between the Rho inhibitors CT03 and CT04?
Answer 1: The only difference between these C3 Transferase proteins (Cat. # CT03 and CT04) is that CT04 is covalently linked to a proprietary cell penetrating moiety via a disulfide bond. In this way, CT04 is a much better reagent to use to inhibit Rho activity in living cells.
Question 2: How can I assess whether Rho activity is changing following CT03 treatment?
Answer 2: There are multiple ways to measure changes in Rho activity. If CT03 has been delivered to cells via micro-injection or pinocytic uptake, we recommend examining Rho-mediated stress fiber formation with fluorescently-labeled phalloidin (Cat. # PHDG1, PHDH1, PHDN1, PHDR1). These Acti-stain phalloidins label F-actin stress fibers. Activation of Rho can be directly quantified with one of our activation assays, either the traditional pull-down (Cat. # BK036) or the RhoA G-LISA activation assay (Cat. # BK124). You can also measure CT03’s ability to ADP ribosylate native Rho in human platelet extracts in vitro (please see the CT03 datasheet for more details). Briefly, a standard biological assay for monitoring the ADP ribosylation of Rho consists of an in vivo ribosylation reaction followed by non-denaturing gel electophoresis and Western blot analysis using Cytoskeleton’s anti-Rho monoclonal antibody (Cat. # ARH03). Stringent quality control ensures that >80% of native Rho protein is ADP ribosylated by the recombinant C3 transferase.
If you have any questions concerning this product, please contact our Technical Service department at tservice@cytoskeleton.com.


