
TECH GUIDE
+3
Loading...
Cat. #BK051/52
Kit contents (96 assays)
Equipment & materials required
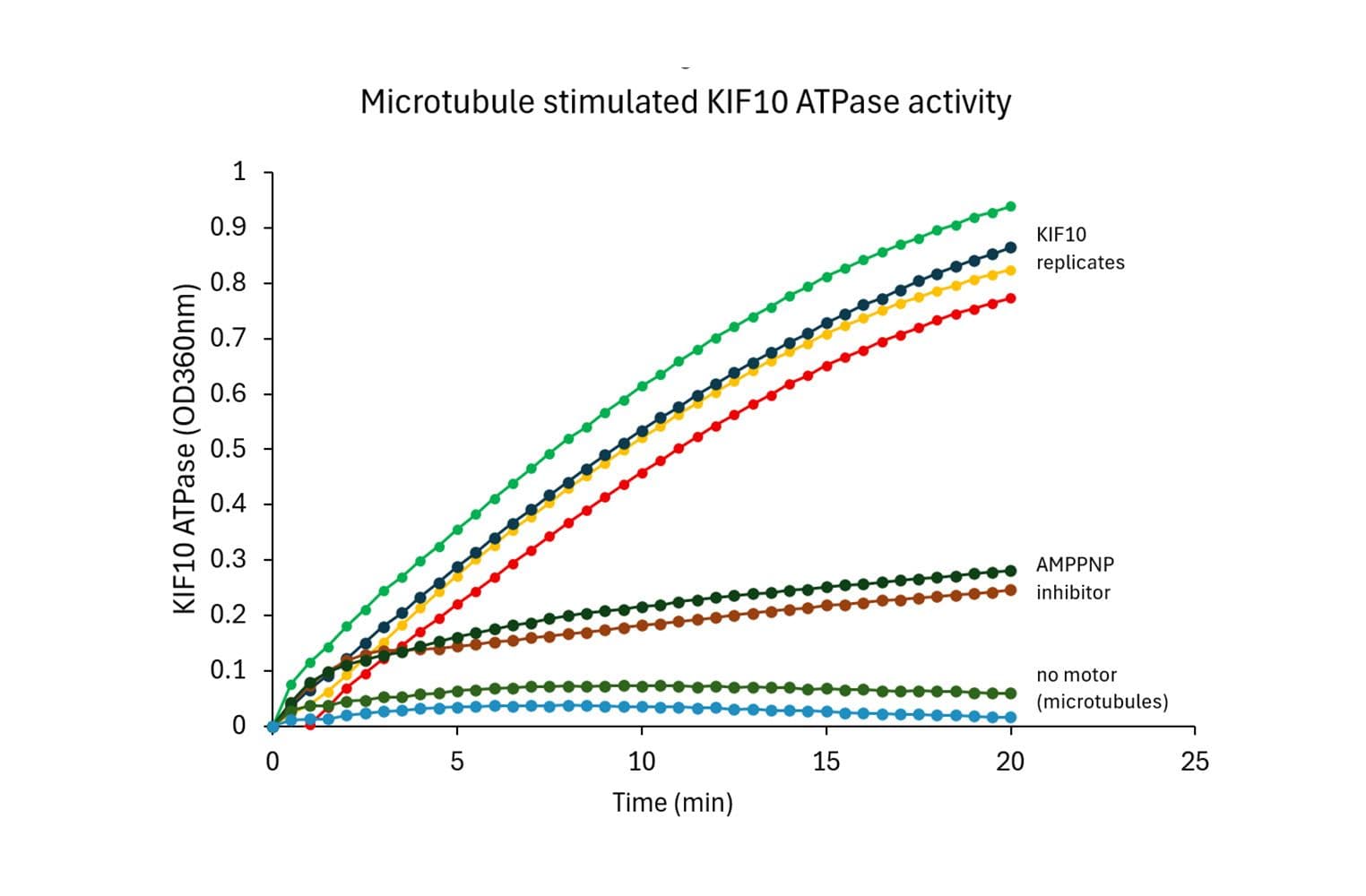
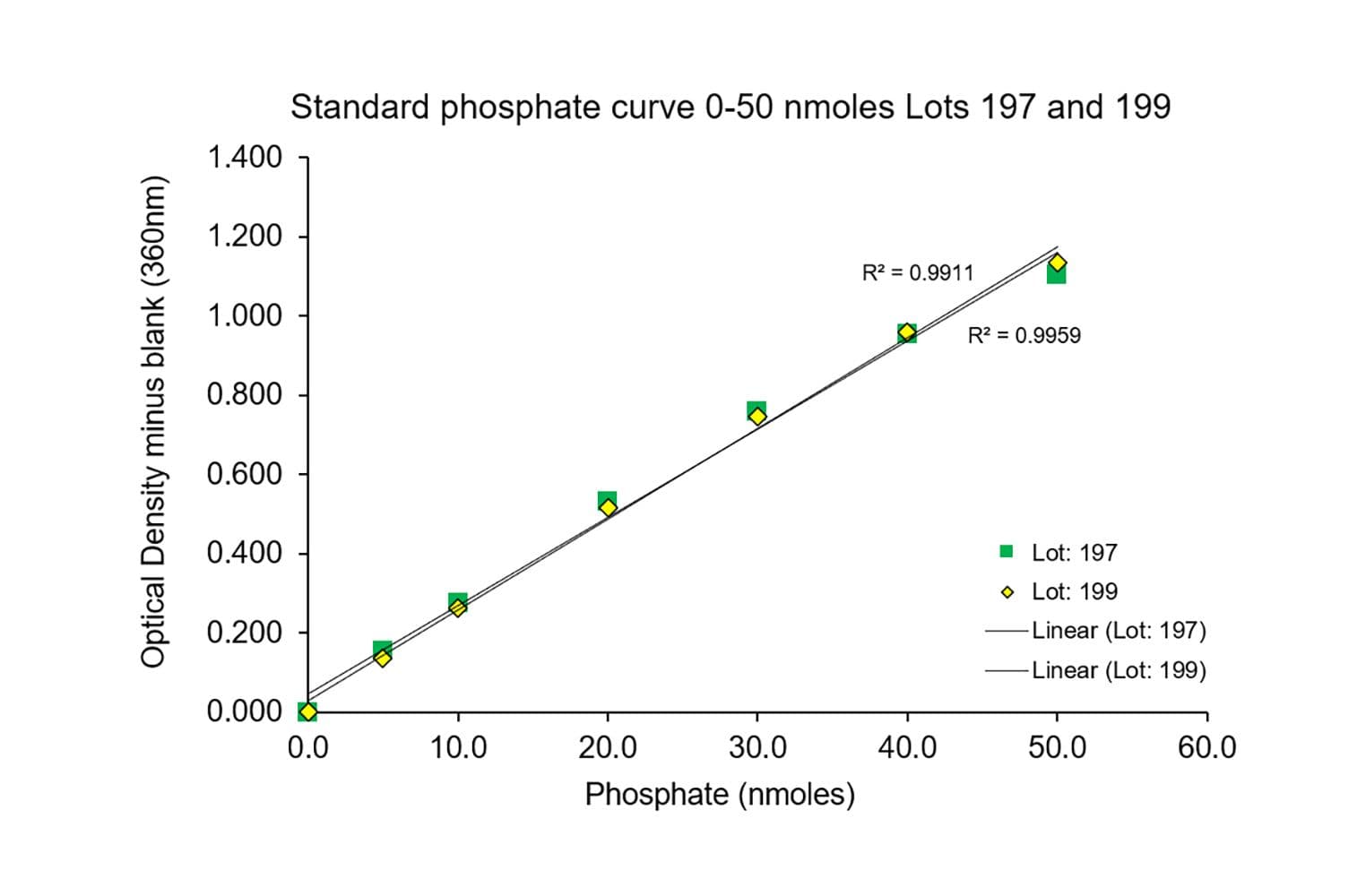
The ELIPA kit quickly measures ATPase and GTPase activity by detecting phosphate (Pi) release. An absorbance change from 330 to 360 nm shows Pi levels, giving fast, accurate results for a wide range of enzymes.
Detection mechanism
The assay is based upon an absorbance shift (330 - 360 nm) that occurs when 2-amino-6-mercapto-7-methylpurine ribonucleoside (MESG) is catalytically converted to 2-amino-6-mercapto-7-methylpurine in the presence of inorganic phosphate (Pi). The reaction is catalyzed by purine nucleoside phosphorylase (PNP). One molecule of inorganic phosphate will yield one molecule of 2-amino-6-mercapto-7-methyl purine in an essentially irreversible reaction. Thus, the absorbance at 360 nm is directly proportional to the amount of Pi generated in the reaction.
Key characteristics






