
TECH GUIDE
+3
Loading...
Kit contents (96 assays: strip-wells allow single or multiple assays per run)
Equipment & materials required
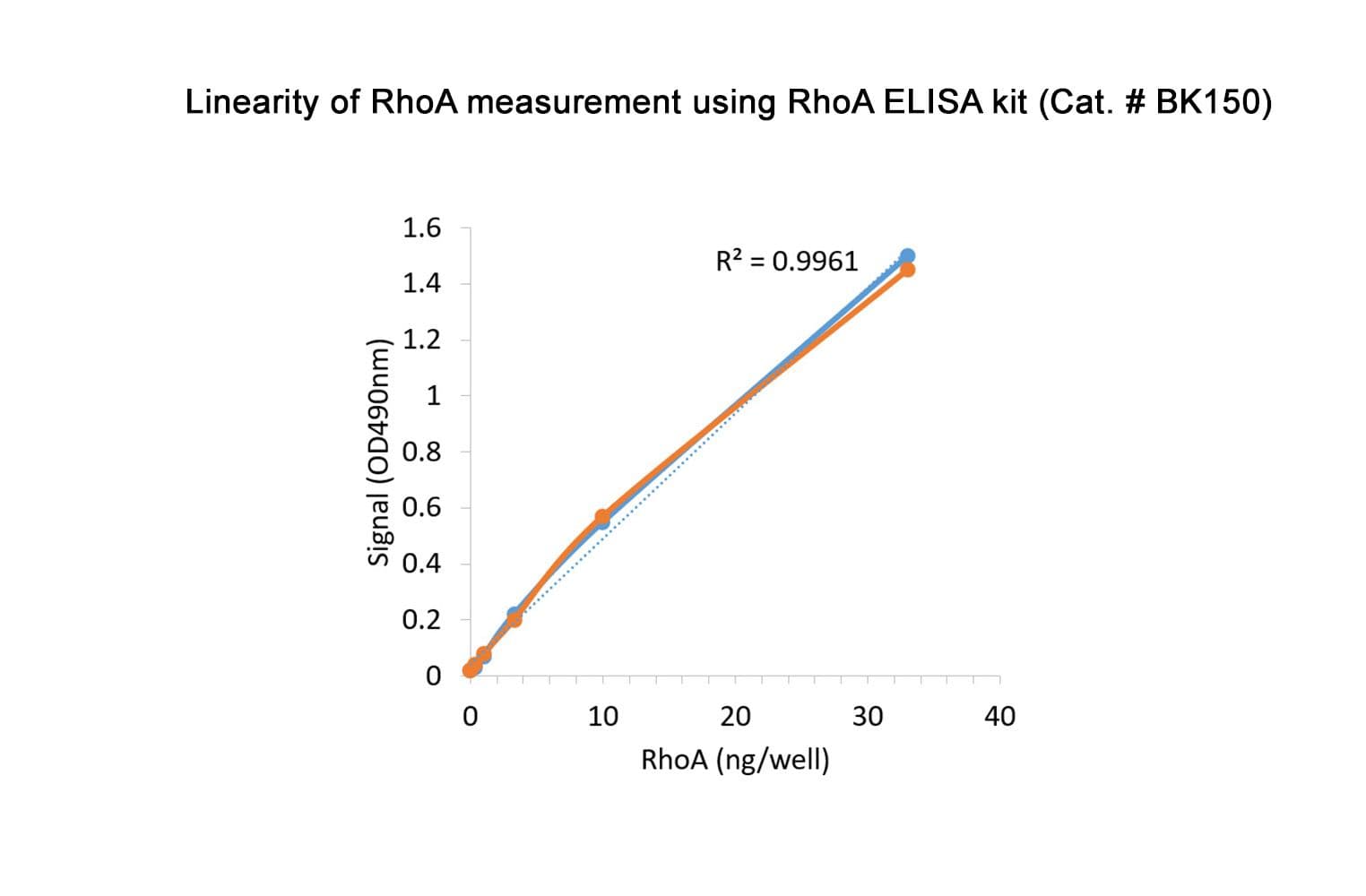
This is a colorimetric ELISA sandwich assay. Each assay well is coated with an anti-Rho IgY antibody, which has a high affinity for RhoA/B/C. A mouse monoclonal detection antibody is specific for RhoA.
Key characteristics






