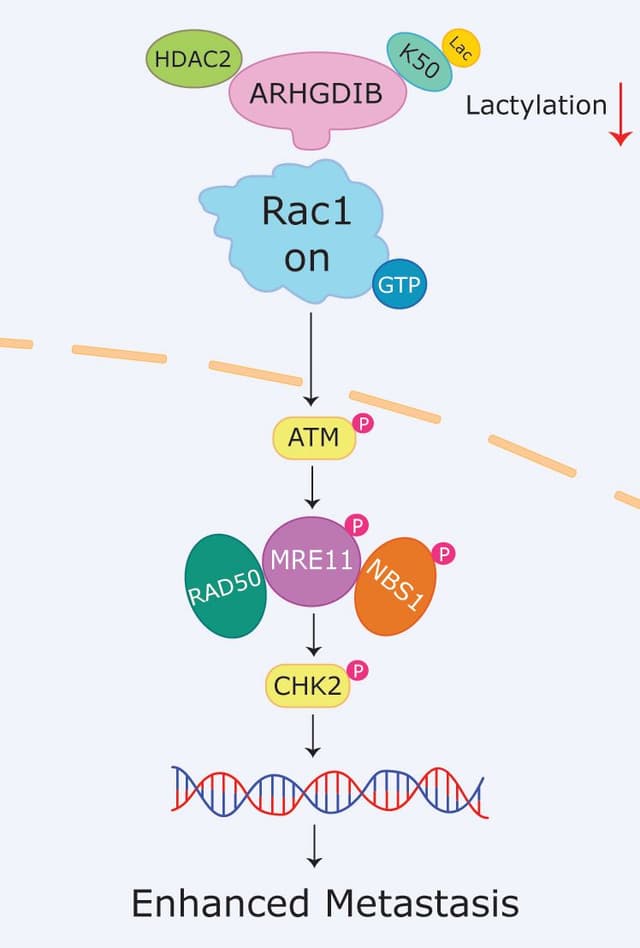
Does Ras regulate spontaneous motility?
Cells have an intrinsic ability to produce random cell migration or basal motility to explore their environment when chemoattractants are not present4. Early studies on Dictyostelium cells lacking Gβ did not activate downstream PI3K signaling events in response to chemoattractants; however, they still produced random motility in the absence of heterotrimeric G proteins19, implying that there may be internal signals that sufficiently control this basal motility. Firtel and colleagues identified the internal signal responsible for basal motility as the Ras-PI3K-F-Actin signaling circuit20. The Devreotes group revealed that Ras and PI3K are the key elements of this excitable network that produces chemotaxis-independent movement in basal motility21. This was supported by Fukushima et al., who showed that an asymmetric distribution of Ras triggered symmetry breaking to produce polarization and directed movement in basal motility22. Furthermore, another study showed that switching between an amoeba-like migratory mode (repeated extending and retracting of pseudopods) to a keratocyte-like gliding (single broad anterior protrusion) could be achieved by altering Ras/Rap-related activities, further implicating Ras as a symmetry-breaking mechanism that regulates migratory mode determination in basal motility23. A current study utilized optogenetic tools to specifically control Ras activity through recruitment of GEFs and GAPs to the front or rear of human neutrophils and showed that this controlled activation of Ras was sufficient to control random motility24. Importantly, this Ras-dependent slow, excitable signaling network does not require the cytoskeleton for activation, but is capable of coupling with cytoskeletal networks to control migration21. Accordingly, recent data suggest that there are complementary feedback loops between this Ras excitable network and increased branch actin polymerization that control front- and back-states of the cell to alter both cell polarity and migration25. Cytoskeleton offer highly sensitive Ras activation and GEF assays to determine the GTP-bound levels of this protein in cell and tissue extracts (see below for more details).
Summary and future insights
These findings collectively point towards a fundamental role for Ras in both chemotactic migration and cell-directed motility. Remarkably, these Ras-dependent signaling events can be activated through external chemoattractants and chemorepellents, or can occur through excitable events, which suggests that the core Ras circuit is essential for cell movement, but must be specifically controlled spatiotemporally through distinct GEFs and GAPs to effectively control these unique types of motile events. For example, what happens to a cell exposed to both a chemoattractant and chemorepellent agent simultaneously? While the current body of work provides key insights towards our understanding of Ras in directed cell movement, there are still more pieces to uncover to fully understand how these different migratory processes converge on Ras to regulate migration.
References
1. Xu, X. and T. Jin, Ras inhibitors gate chemoattractant concentration range for chemotaxis through controlling GPCR-mediated adaptation and cell sensitivity. Front Immunol, 2022. 13: p. 1020117.
2. Van Haastert, P.J. and P.N. Devreotes, Chemotaxis: signalling the way forward. Nat Rev Mol Cell Biol, 2004. 5(8): p. 626-34.
3. Metzemaekers, M., M. Gouwy, and P. Proost, Neutrophil chemoattractant receptors in health and disease: double-edged swords. Cell Mol Immunol, 2020. 17(5): p. 433-450.
4. Matsuoka, S., et al., Spontaneous signal generation by an excitable system for cell migration. Front Cell Dev Biol, 2024. 12: p. 1373609.
5. Jin, T., et al., Localization of the G protein betagamma complex in living cells during chemotaxis. Science, 2000. 287(5455): p. 1034-6.
6. Sasaki, A.T., et al., Localized Ras signaling at the leading edge regulates PI3K, cell polarity, and directional cell movement. J Cell Biol, 2004. 167(3): p. 505-18.
7. Kae, H., et al., Chemoattractant-induced Ras activation during Dictyostelium aggregation. EMBO Rep, 2004. 5(6): p. 602-6.
8. Kortholt, A., et al., Dictyostelium chemotaxis: essential Ras activation and accessory signalling pathways for amplification. EMBO Rep, 2011. 12(12): p. 1273-9.
9. Kortholt, A., et al., Ras activation and symmetry breaking during Dictyostelium chemotaxis. J Cell Sci, 2013. 126(Pt 19): p. 4502-13.
10. Liu, Y., et al., A Galpha-Stimulated RapGEF Is a Receptor-Proximal Regulator of Dictyostelium Chemotaxis. Dev Cell, 2016. 37(5): p. 458-72.
11. Lin, Y., et al., Ras-mediated dynamic and biphasic regulation of cell migration. Proc Natl Acad Sci U S A, 2025. 122(30): p. e2503847122.
12. Lin, Y., et al., Ras suppression potentiates rear actomyosin contractility-driven cell polarization and migration. Nat Cell Biol, 2024. 26(7): p. 1062-1076.
13. Xu, X., et al., GPCR-controlled membrane recruitment of negative regulator C2GAP1 locally inhibits Ras signaling for adaptation and long-range chemotaxis. Proc Natl Acad Sci U S A, 2017. 114(47): p. E10092-E10101.
14. Phillips, J.E. and R.H. Gomer, A secreted protein is an endogenous chemorepellant in Dictyostelium discoideum. Proc Natl Acad Sci U S A, 2012. 109(27): p. 10990-5.
15. Rijal, R., et al., An endogenous chemorepellent directs cell movement by inhibiting pseudopods at one side of cells. Mol Biol Cell, 2019. 30(2): p. 242-255.
16. Kirolos, S.A. and R.H. Gomer, A chemorepellent inhibits local Ras activation to inhibit pseudopod formation to bias cell movement away from the chemorepellent. Mol Biol Cell, 2022. 33(1): p. ar9.
17. Kirolos, S.A., et al., A phosphatidylinositol phosphate kinase inhibits Ras activation and regulates chemorepulsion in Dictyostelium discoideum. J Cell Sci, 2023. 136(14).
18. El-Sobky, M.H., R. Rijal, and R.H. Gomer, Two endogenous Dictyostelium discoideum chemorepellents use different mechanisms to induce repulsion. Proc Natl Acad Sci U S A, 2025. 122(22): p. e2503168122.
19. Wu, L., et al., The G protein beta subunit is essential for multiple responses to chemoattractants in Dictyostelium. J Cell Biol, 1995. 129(6): p. 1667-75.
20. Sasaki, A.T., et al., G protein-independent Ras/PI3K/F-actin circuit regulates basic cell motility. J Cell Biol, 2007. 178(2): p. 185-91.
21. Huang, C.H., et al., An excitable signal integrator couples to an idling cytoskeletal oscillator to drive cell migration. Nat Cell Biol, 2013. 15(11): p. 1307-16.
22. Fukushima, S., S. Matsuoka, and M. Ueda, Excitable dynamics of Ras triggers spontaneous symmetry breaking of PIP3 signaling in motile cells. J Cell Sci, 2019. 132(5).
23. Miao, Y., et al., Altering the threshold of an excitable signal transduction network changes cell migratory modes. Nat Cell Biol, 2017. 19(4): p. 329-340.
24. Pal, D.S., et al., Actuation of single downstream nodes in growth factor network steers immune cell migration. Dev Cell, 2023. 58(13): p. 1170-1188 e7.
25. Kuhn, J., et al., Complementary cytoskeletal feedback loops control signal transduction excitability and cell polarity. Nat Commun, 2025. 16(1): p. 7482.