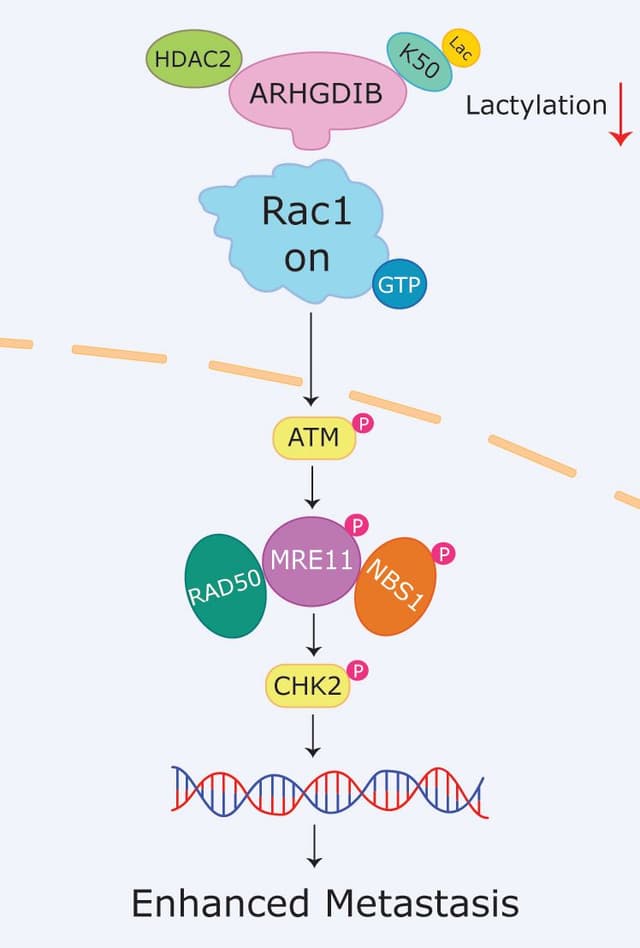
Is there more to learn about Cdc42 in the gut?
The overall body of work describes a critical role for Cdc42 in the physiologic and pathologic function of the gastrointestinal system. While intriguing studies highlight Cdc42’s importance in host cell and microbiota crosstalk for immune cell regulation under physiologic conditions, it will be interesting to see if it also plays a similar role in IBD, gastric cancer, and other gastrointestinal diseases.
Furthermore, it will be fascinating to determine which of the mechanistic functions and regulatory pathways described above are most critical for Cdc42’s impact on gastric health and disease, as these could have profound implications for therapeutic intervention.
Check out Cytoskeleton's Cdc42 Biochem kit here, in addition to our Cdc42 learning resources here.
References
1. Danne, C., et al., Neutrophils: from IBD to the gut microbiota. Nat Rev Gastroenterol Hepatol, 2024. 21(3): p. 184-197.
2. Zundler, S., et al., Gut immune cell trafficking: inter-organ communication and immune-mediated inflammation. Nat Rev Gastroenterol Hepatol, 2023. 20(1): p. 50-64.
3. Wong, C.C. and J. Yu, Gut microbiota in colorectal cancer development and therapy. Nat Rev Clin Oncol, 2023. 20(7): p. 429-452.
4. Kaur, S., et al., Potential biomarkers in early detection of gastric cancer. Front Pharmacol, 2025. 16: p. 1642927.
5. Fu, J., et al., The role of cell division control protein 42 in tumor and non-tumor diseases: A systematic review. J Cancer, 2022. 13(3): p. 800-814.
6. Yasumi, T., CDC42 missense mutations and human diseases: from neurodevelopmental disorders to autoinflammation. J Biochem, 2025. 178(2): p. 73-78.
7. Ladinsky, M.S., et al., Endocytosis of commensal antigens by intestinal epithelial cells regulates mucosal T cell homeostasis. Science, 2019. 363(6431).
8. Melendez, J., et al., Cdc42 coordinates proliferation, polarity, migration, and differentiation of small intestinal epithelial cells in mice. Gastroenterology, 2013. 145(4): p. 808-19.
9. Sakamori, R., et al., Cdc42 and Rab8a are critical for intestinal stem cell division, survival, and differentiation in mice. J Clin Invest, 2012. 122(3): p. 1052-65.
10. Zhang, Z., et al., CDC42 controlled apical-basal polarity regulates intestinal stem cell to transit amplifying cell fate transition via YAP-EGF-mTOR signaling. Cell Rep, 2022. 38(2): p. 110009.
11. Zhang, D., et al., Monogenic deficiency in murine intestinal Cdc42 leads to mucosal inflammation that induces crypt dysplasia. Genes Dis, 2024. 11(1): p. 413-429.
12. Cho, H.S., et al., Structure of gut microbial glycolipid modulates host inflammatory response. Cell, 2025. 188(19): p. 5295-5312 e18.
13. Li, X., et al., Role of Rho GTPases in inflammatory bowel disease. Cell Death Discov, 2023. 9(1): p. 24.
14. Dong, L.M., et al., Cell division cycle protein 42 regulates the inflammatory response in mice bearing inflammatory bowel disease. Artif Cells Nanomed Biotechnol, 2019. 47(1): p. 1833-1838.
15. Liu, L., et al., Cell division control 42 elevates during infliximab therapy, and its increment relates to treatment response in ulcerative colitis patients. J Clin Lab Anal, 2022. 36(6): p. e24477.
16. Hu, Q., et al., Paeoniflorin alleviates DSS-induced ulcerative colitis by suppressing inflammation, oxidative stress, and apoptosis via regulating serum metabolites and inhibiting CDC42/JNK signaling pathway. Int Immunopharmacol, 2024. 142(Pt A): p. 113039.
17. Stojanovic, M., et al., Hyperglycemia alters the gene and protein expression of CDC42 in small and large intestine of Sprague-Dawley rats. Mol Cell Biochem, 2026.
18. Wang, D.N., et al., Bacterial infection promotes tumorigenesis of colorectal cancer via regulating CDC42 acetylation. PLoS Pathog, 2023. 19(2): p. e1011189.
19. Liu, M., et al., miR-137 targets Cdc42 expression, induces cell cycle G1 arrest and inhibits invasion in colorectal cancer cells. Int J Cancer, 2011. 128(6): p. 1269-79.
20. Liu, M., et al., miR-185 targets RhoA and Cdc42 expression and inhibits the proliferation potential of human colorectal cells. Cancer Lett, 2011. 301(2): p. 151-60.
21. Liang, X., et al., Depletion of LINC01133 from cancer-associated fibroblasts exacerbates the progression of gastric adenocarcinoma. iScience, 2025. 28(12): p. 113886.
22. Gomez Del Pulgar, T., et al., Cdc42 is highly expressed in colorectal adenocarcinoma and downregulates ID4 through an epigenetic mechanism. Int J Oncol, 2008. 33(1): p. 185-93.
23. Al-Tassan, N.A., et al., A new GWAS and meta-analysis with 1000Genomes imputation identifies novel risk variants for colorectal cancer. Sci Rep, 2015. 5: p. 10442.
24. Fan, H., et al., Urine proteomic signatures predicting the progression from premalignancy to malignant gastric cancer. EBioMedicine, 2022. 86: p. 104340.
25. Sakamori, R., et al., CDC42 inhibition suppresses progression of incipient intestinal tumors. Cancer Res, 2014. 74(19): p. 5480-92.
26. Valdes-Mora, F., et al., Clinical relevance of the transcriptional signature regulated by CDC42 in colorectal cancer. Oncotarget, 2017. 8(16): p. 26755-26770.
27. Ma, L.L., et al., Cdc42 subcellular relocation in response to VEGF/NRP1 engagement is associated with the poor prognosis of colorectal cancer. Cell Death Dis, 2020. 11(3): p. 171.
28. Du, D.S., et al., Effects of CDC42 on the proliferation and invasion of gastric cancer cells. Mol Med Rep, 2016. 13(1): p. 550-4.
29. Wang, Q., et al., MICAL2 contributes to gastric cancer cell migration via Cdc42-dependent activation of E-cadherin/beta-catenin signaling pathway. Cell Commun Signal, 2022. 20(1): p. 136.