
TECH GUIDE
+3
Loading...
Cat. #T240
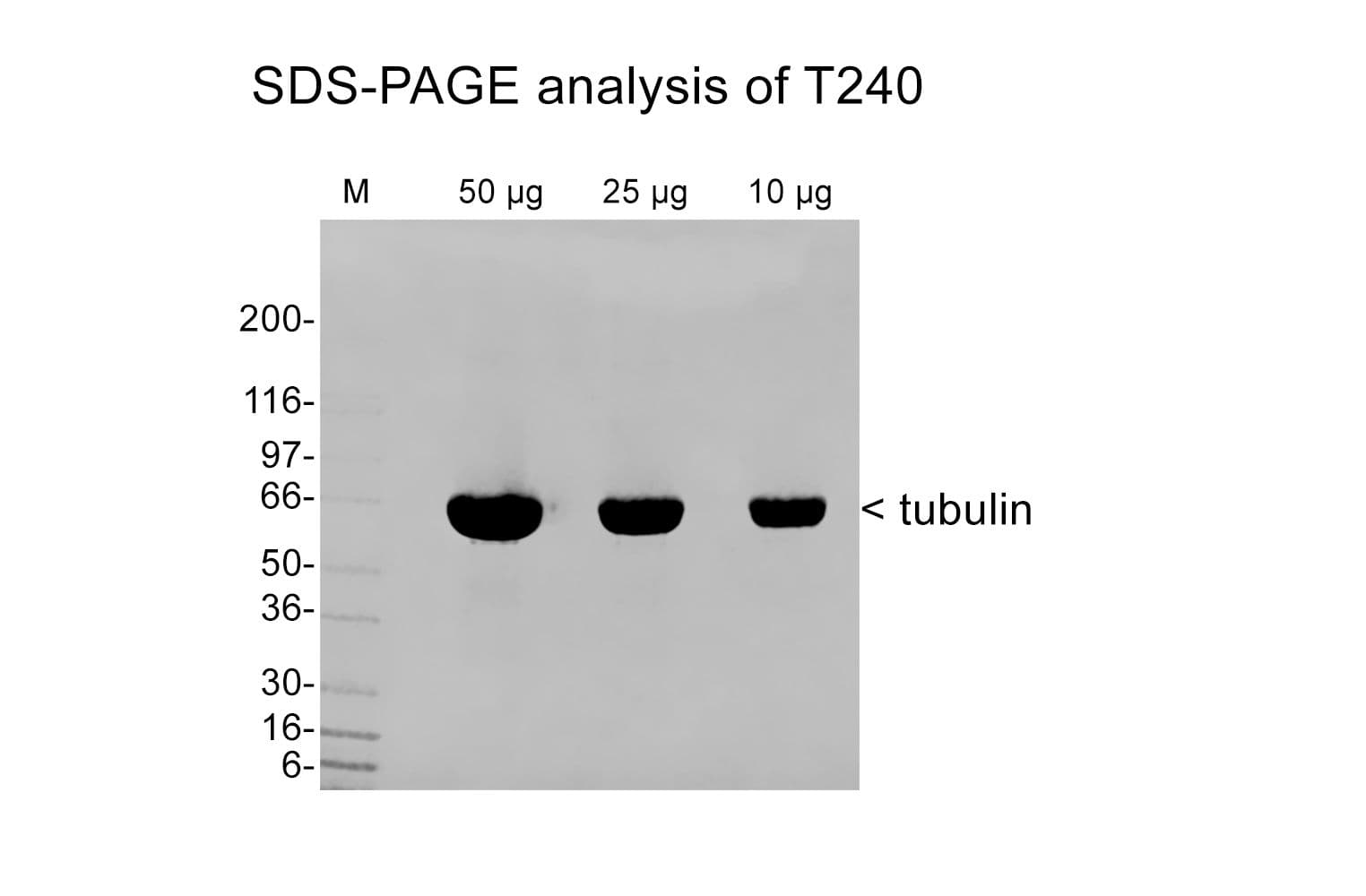
Tubulin is purified from porcine brain using a modified Shelanski et al. method, followed by cation exchange chromatography to achieve >99% purity. Supplied as a lyophilized powder. Shipped at ambient temperature, powder stored at 4°C.
Tubulin is a heterodimer of α- and β-isotypes, each 55 kDa. SDS-PAGE shows a single 55 kDa band. The molecular weight of the functional heterodimer is 110 kDa.
Protein purity is assessed by scanning densitometry of Coomassie Blue stained protein on a12% polyacrylamide gel. Purity is determined to be >99% pure.
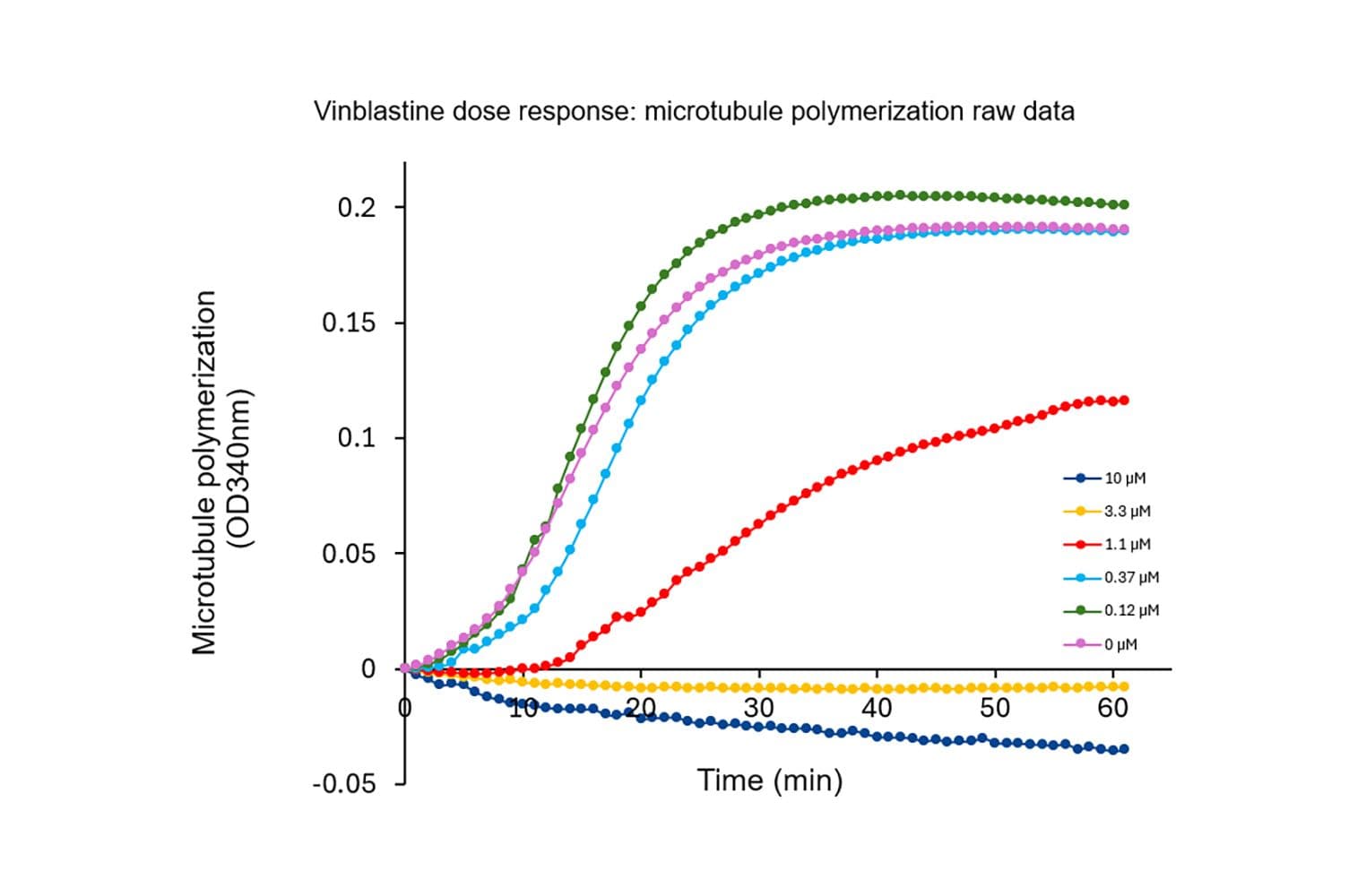
The biological activity of T240 is evaluated using a tubulin polymerization assay, which monitors microtubule formation by measuring absorbance at 340 nm. Under the test conditions (5 mg/ml tubulin in General Tubulin Buffer with 5% glycerol and 1 mM GTP, 180 µl volume, 37 °C, 0.8 cm pathlength), the OD340nm should reach between 0.78 and 1.1 within 30 minutes.







