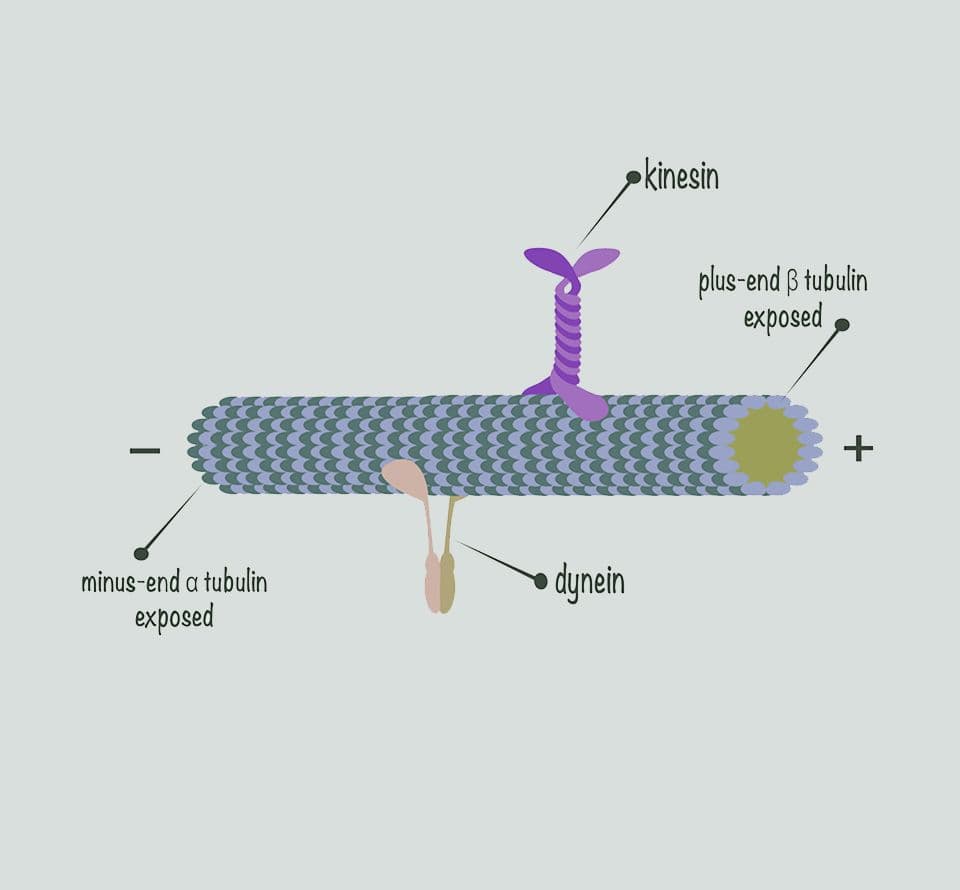
Lab-side feedback: kinesin motor assay
Cytoskeleton’s tech support was able to answer every single question, no matter how oddly specific it was. It was clear that they’re really familiar with the product and can help us work through any experimental challenges.”

Zoe Kaegi
Graduate student
University of Miami